
Picking up on some of the BIG IDEAS in brain research, which was wonderfully chaotic when we last discussed in December under the hashtag #BrainIdeasCountdown, e.g. https://neuromatch.social/@NicoleCRust/109557289393362842
Here's an attempt to fill in some blanks, and let's flip the hashtag: #BigBrainIdeas. I'll focus on the notion that there are facts, ideas and then there are "Big Ideas" and I'll focus on the last one. Please join in!
I'd argue that one of the most influential Big Ideas about the brain in the latter half of the 20th century is the is the notion that:
The neocortex of the brain is made up of a generic functional element that is repeated again and again and from this repetition, all of cortical function emerges
I'm talking about the cortical column, first described by Vernon Mountcastle in 1957. The unit contains ~10K neurons and humans have ~25 million of them. The rapid evolution of humans is proposed to have followed from a rapid expansion of cortex that happened because it has this repetitive crystalline structure. The gist behind the "functional" bit is that each unit always does the same generic computation, and the different functions of different brain areas result from the different inputs that these units receive. @TrackingActions very nicely summarizes the ideas here: https://www.nature.com/articles/s41583-022-00658-6
So what does this generic functional unit do? Proposals vary. One idea, also reflected in deep convolutional neural networks, is that it does two(ish) things: selectivity and invariance, stacked repetitively to support things like recognizing objects. Other proposals suggest that the brain is a prediction machine and each unit contributes a little bit to those predictions in a manner that relies not just on feedforward connectivity, but also feedback. Some proposals suggest that the function of the unit varies along a gradient as a consequence of biophysical properties like receptor expression: https://www.nature.com/articles/s41583-020-0262-x.
Among brain researchers, this Big Idea is polarizing - obvious to some and misguided to others. Where are you in terms of your 'buy in' with this big idea?

@NicoleCRust @TrackingActions @cogneurophys I am not sure that this idea has actually been very influential on the grand scale of neurocog theories. The modern theoretical approach seems to revolve around understanding how brain areas connect in networks to solve problems, and I can't see how generic computation would inspire/drive this perspective.
@bwyble @TrackingActions @cogneurophys
The idea spawned maps like this, which have been highly influential for neurocog theories, no?
@NicoleCRust @TrackingActions @cogneurophys I thought that diagram was the result of neuroanatomy studies. Why would a generalized function theory lead to a highly specific wiring diagram like this?
@bwyble
Yes, but ... Felleman & Van Essen defined the hierarchical levels of this diagram according to the cannonical microcircuit rule: L4 receives input; L2/3=feedforward output; L4/5=feedback output.

That rule was derived from anatomy too.
@DrYohanJohn @NicoleCRust I agree with Yohan, I don't think the microcircuit is crucial there, rather it's observing that there are laminar patterns that are more or less ubiquitous.
@bwyble @DrYohanJohn
Interesting! To me, those ideas could not be more connected.

@NicoleCRust @bwyble @DrYohanJohn
shameless plug, read my CONB on this topic with Adam K, for my more edited, fleshed out opinion!
https://www.sciencedirect.com/science/article/pii/S0959438822001246
(just insert "microcircuit motif" whereever you read cell-type, Adam K. and I disagree about which phrase is the better one :p)
I get the argument about confusing implementation and computation, but I agree with with @NicoleCRust that the idea of canonical cortical computations has been super influential, especially / at least in vision (which is all of computational neuro anyway, amiright?)
I think the idea of simple, repeated computation is kind of necessary / permissive for a certain types of "grand unified theory" that are very intuitively appealing exactly because they squash together computation and implementation into one little thing that is intuitively understandable in words. The fact that these "theories" are conceptually "small" down to implementation mean that people understand them, they catch on, they drive research.
Now whether this is positive or not, is an open debate. But I think there's no way we can say that ideas like predictive processing, backprop, divisive normalization, and maybe even convolution havent been wildly influential in the field.
To state this in a slightly more aggressive way than I feel: I do think the idea of a canonical microcircuit is very useful, because it's studiable! If we just start off by assuming everything is brain soup it's very easy to just give up and assume we'll never understand implementation. I rather start off with the assumption that there exist architectural motifs that matter, and try to take that as a hypothesis from which to start, than just admit defeat.
So maybe the answer is, it depends on the level of explanation you are lookign for. If you dont care baout multilevel understanding then "how brain regions connect in networks" black box type of understanding is enough. I personally think it's only step 1, and then understanding more details of implementation is step 2. Furthermore, I've come to think that the implementation detaiuls will probably constrain and help us understand the higher level.

@achristensen56 @NicoleCRust @bwyble @DrYohanJohn
The fact that, when, say, sight is lost early, other sensory modalities can successfully invade areas they wouldn't necessarily be processed, in suggests standardisation of function/microanatomy across large parts of the brain.

@strangetruther @achristensen56 @NicoleCRust @bwyble @DrYohanJohn
A simple argument for some sort of canonical cortical computation is: "cerebral cortex ... processes ... diverse tasks with what appears to be a remarkably uniform, primarily six-layer architecture, albeit with significant differences in details across species and cortical areas [1,2,3,4–10,11,12–14]. ... This has long suggested the idea that a piece of six-layer cortex with a surface area on the order of a square millimeter constitutes a fundamental cortical ‘processing unit’ [e.g. 16,17].The cortex varies in surface area by a factor of 10000 across a set of 37 mammalian species, while thickness (the distance across the layers) varies only by a factor of 10 over the same species [18], suggesting that the most salient evolutionary change in cortex has been enormous multiplication of the number of ‘units’ [e.g. 14]."
The last two references are:
14. Rakic P: Confusing cortical columns. Proc Natl Acad Sci U S A 2008, 105:12099-12100.
18. Hofman MA: On the evolution and geometry of the brain in mammals. Prog Neurobiol 1989, 32:137-158.
This is from the 1st paragraph of a Current Opinion review paper that I wrote that Nicole cited further up in the thread (https://pubmed.ncbi.nlm.nih.gov/26868041/)
More generally I think there are (at least) 4 mammalian (and, except for cortex, vertebrate) brain structures that each clearly have repeating architecture, and that -- at least as studied in primates -- communicate pretty intimately with one another: cortex, thalamus, basal ganglia, and cerebellum. They communicate with specificity, eg a given piece of cortex communicates with given thalamic nuclei and given regions of basal ganglia and cerebellum, which communicate with one another, e.g. Boston & Strick https://www.nature.com/articles/s41583-018-0002-7. These specific ctx/BG/cerebellum interactions cover at least posterior parietal through frontal cortex, and perhaps higher sensory cortices as well, i.e. they cover all sorts of cognitive processing, not just motor processing which is the traditional function assigned to BG and cerebellum. So it's not just the enormous multiplication of cortical "units" (with diversification, i.e. the spectrum Y.J. referred to) , but also the corresponding multiplication of their partner thalamic, basal ganglia, and cerebellar "units" that suggest some fundamental computional operation, albeit again with diversification.
You don't see this sort of thing in the brainstem. Different bespoke nuclei or other sorts of neural units each do different pieces of different computations. In contrast, the existence of these structures with repeated modular subunits with roughly repeating architectures (despite much variability and diversification), and with specific patterns of interconnections between their modules, as well as their enormous growth in numbers of modules at least in mammalian evolution, all just scream out that some sort of computational motif is being repeated (with variations on the repeated units, much as multiple copies of a gene provide a substrate for evolution into multiple variants -- and occasionally quite new structures). That wouldn't happen by accident.
Kenji Doya long ago postulated that cortex is for associative learning, BG for reinforcement learning, and cerebellum for error-correcting learning. That still sounds like a decent 0th-order take. And, I'll add my speculation, one function of thalamus -- not all that it is doing -- is to take any modality of information whatsoever and convert it into a language that cortex understands, using a roughly uniform architecture with roughly uniform biophysics across all these different modalities of information.
@bwyble @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
Well, the question is, what is the level of the unit that you see repeated? The fact that individual neurons are repeated would not account for any of the specificity of the repeated architectures one sees across each of the four structures. Based on about a 1sq mm chunk of V1 containing all preferred orientations from both eyes at a given retinotopic position, with the next chunk over (e.g., about 1mm away) having half-overlapping receptive fields, Hubel and Wiesel suggested that size of chunk as a cortical processing unit (the size varies across species from 1/2 mm to 1.5 mm or so, but order of magnitude 1 mm). That is at least roughly consistent with dense connectivity in cortex extending horizontally maybe 200-300 microns from any given point, along with sparser connectivity over longer distances. Comparable structures have been seen in many other cortical areas. So that is what I think of as the unit that repeats. Others have focused on a 25 x 25 micron or so "minicolumn", or the area spanned by the neurons spawned from a given radial glial cell. That also repeats, but personally seems to me too small to be a computational unit, i.e. to do a self-contained computation.
But more generally, I'd want to characterize the repeated units of each of the four structures that are connected, that talk to one another -- a thalamo-cortical-basal-ganglia-cerebellar unit. I suspect the cortical part is something like Hubel & Wiesel's square mm, but maybe it would be more like a cortical area, I'm really not sure.
@jiahongbo @bwyble @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
I'm sure orientation and ocular dominance do not begin to describe what V1 is doing. But they are two important and prominent things that V1 represents, and so a region that gives a complete local representation of them is likely to have a complete local representation of the visual scene.

@kendmiller @bwyble @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn yeah but then it's not V1 of the mouse or monkey or human subject, but rather the manifolds in digital computers doing what people assume V1 does...
@kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
Yes, I think that's the hard/interesting question about the scale of replication. Perhaps one can make a fairly strong inference about V1 from observations of orientation sensitivity and also ocular dominance, but that might be specific to V1. In my view, the brain likes to specialize as much as it likes to repeat, and the two demands play off against each other.
What I'd like to see to be more confident of repeating columns is evidence somewhere else, like tonotopy, or striping in the motor system. That might exist but I've never heard of it.

@bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
Mixed selectivity multifunctional neurons seem to be common in cortex. That argues for a lot of repetition and not a lot of strict specialization.
@ekmiller @kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
They do, but there's also a lot of ways that neural tissue specializes as well, especially when one looks at areas like the hippocampus.
I've always been a splitter rather than a lumper though, and this has affected my perspective on everything neuro.
@bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
I'm more of a splitter when it comes to the brain's infrastructure. But anatomy is like the road-and-highway system. It says where traffic *could* go, not where it flows from moment to moment.
Cortical function, IMHO, will be about emergent properties and highly interactive dynamics that have less respective for boundaries and splits.
That is not to say that infrastructure isn't important. It's foundational
@ekmiller @bwyble @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
Brad, re evidence of repeating columns outside V1: you see it in V2, MT, V4, IT. More generally, a sign of periodic organization of preferred features is the patchy arrangement of long-range (>1mm) horizontal projections, which in V1 are patchy because they're targeting other patches of similar preferred orientation. My memory of old anatomy papers on higher areas of monkey cortex are that, wherever you look in primate cortex, you see such patchiness of long-range connections. I've forgotten what papers those were though, quite possibly from Jenny Lund, or maybe Kathy Rockland, or someone else ...
@kendmiller @ekmiller @bwyble @strangetruther @achristensen56
@DrYohanJohn
What an amazing thread ⬆️!

@NicoleCRust u have a knack for making these :)

@NicoleCRust @kendmiller @ekmiller @bwyble @strangetruther @achristensen56 @DrYohanJohn
About "what cortex does" and the repeating unit idea, I wanted to encourage more of an evolutionary perspective.
No doubt that the mammalian cortex is a beautiful invention of nature, but in understanding the "macro modules" as Ken suggested (cortex, basal ganglia, cerebellum, thalamus, if I recall correctly) and what they might do as a sort of unit, we should recall that cortex is (most of) the mammalian pallium (basolateral amygdala also has pallial origins).
And if we look at the vertebrate pallium in general it definitely is not uniform and has at least 4-6 domains that are very important to separate if we want to understand homology across vertebrates. They are uninspiredly called lateral, medial, ventral, and dorsal.
For example, medial pallium is linked to the hippocampus, and is involved in navigation across amniotes (at least).
Also important is that the macro-architecture linking pallium, thalamus, and basal ganglia is a theme seen across amniotes (at least), not just mammals. Here's a figure of how we summarized this (together with comparative neuroanatomists who really know what they are talking about; I'm just a fan).
@NicoleCRust @kendmiller @ekmiller @bwyble @strangetruther @achristensen56 @DrYohanJohn
Here's another figure that is useful. Of course all of this is a working model, but there's quite a lot of work from the "Spanish school" that supports these ideas, although my colleagues in particular are now favoring a model of pallium that has 6 domains instead of 4.
In any case, these evolutionary considerations seem key to "understanding the brain" but not really very en vogue these days, even more so in the US. Some excellent work is done in Europe however, probably given the funding structure.
@PessoaBrain @NicoleCRust @kendmiller @ekmiller @bwyble @achristensen56 @DrYohanJohn
Intriguing!! Thanks!
@PessoaBrain @NicoleCRust @kendmiller @bwyble @strangetruther @achristensen56 @DrYohanJohn
Nice. Thanks.
I like to say that the cortex is an update to the subcortex. It elaborated on the functions the primitive brain areas were already doing.
One of those added features is integration.
Tying into Luiz's point, the cortex and the basal ganglia expanded in tandem.
"With the appearance of neocortex in mammals, however, basal ganglia outputs to motor cortex via thalamus became of greater significance, especially in primates, in which a parallel expansion of cerebral cortex and basal ganglia occurred."
https://link.springer.com/chapter/10.1007/978-1-4419-0340-2_1
@ekmiller @PessoaBrain @NicoleCRust @kendmiller @bwyble @strangetruther @achristensen56
@DrYohanJohn @PessoaBrain @NicoleCRust @kendmiller @bwyble @strangetruther @achristensen56
+1 You can't understand cortex without the subcortex.
Same goes for the BG. Cognition evolved from the volitional control of movement.
We tried to capture that here:
Rules through Recursion: How Interactions between the Frontal Cortex and Basal Ganglia May Build Abstract, Complex Rules from Concrete, Simple Ones
https://collaborate.princeton.edu/en/publications/rules-through-recursion-how-interactions-between-the-frontal-cort
@DrYohanJohn @PessoaBrain @NicoleCRust @kendmiller @bwyble @strangetruther @achristensen56
Here's one with a cheekier title.
Bootstrapping your brain: How interactions between the frontal cortex and basal ganglia may produce organized actions and lofty thoughts
https://doi.org/10.1016/B978-012372540-0/50011-X
Clearly, this was all before my self-imposed ban of using colons (:) in titles.
@ekmiller @DrYohanJohn @PessoaBrain @NicoleCRust @kendmiller @bwyble @achristensen56
Colons not so much a problem on a Mac- when saving such items to disc, Macs seem to remove the colons! (I still like colons in titles!).
On your papers (I could only access the abstracts), you have the familiar "averaged out, and non-goal" nature of representing associations in the cortex, and in the BG, indiv episodes, PLUS more goal centred.
Are "episodes" (Hip.cs) and "goal centred e.g. DA" separate?
@strangetruther @DrYohanJohn @PessoaBrain @NicoleCRust @kendmiller @bwyble @achristensen56
Thanks.
A PDF of Rules Through Recursion is available here:
https://ekmillerlab.mit.edu/publications/
@ekmiller @DrYohanJohn @PessoaBrain @NicoleCRust @kendmiller @bwyble @achristensen56
Ooh! Very interesting! Thanks so much EM!
I'm (finally!) implementing construction of higher rules etc from lower ones in my system.
It would be so nice to locate these in wet ware!!
Nice we've got birds to cf.
Their brains got bigger when mammal's did: constant heat allowed bodies suited to it, but when the heat eased, it had to be generated. As energy for brains made heat, energy to run them was now free!
@ekmiller @DrYohanJohn @NicoleCRust @kendmiller @bwyble @strangetruther @achristensen56
Cool, will take a look at the paper and use it in future reviews.
Even cooler, the bird brain is crazy integrative too. It has loops via basal ganglia via dorsal and ventral striatum, and repeats many themes of the mammalian organization.

Add cerebellum and make it a threesome. Here's a WM paper that mentions cerebellum 74 times... gotta like that.
https://pubmed.ncbi.nlm.nih.gov/31039359/
@DrYohanJohn @ekmiller @PessoaBrain @NicoleCRust @kendmiller @bwyble @strangetruther @achristensen56
For sure. I remember my surprise by this result.
We report that persistent representation of information in frontal cortex during motor planning is dependent on the cerebellum.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6212318/
A cortico-cerebellar loop for motor planning
@dsmith @DrYohanJohn @ekmiller @PessoaBrain @kendmiller @bwyble @strangetruther @achristensen56
Nice!
How does fit into the cerebellar automaticity story, @macshine ?
[I'm referring to this: https://academic.oup.com/brain/article-abstract/145/9/2967/6648879?redirectedFrom=fulltext&login=true ]
@NicoleCRust @dsmith @ekmiller @PessoaBrain @kendmiller @bwyble @strangetruther @achristensen56

@DrYohanJohn @NicoleCRust @dsmith @ekmiller @PessoaBrain @kendmiller @bwyble @strangetruther @achristensen56 IMHO it fits beautifully -- so much of our cognitive capacity comes down to our ability to rapidly anticipate possible next steps in complex sequences of behaviour, which the cerebellum (via its connections through the pons + thalamus) are ideally suited to supply back to the frontal cortex as a part of dynamic, adaptive cognitive processes
@macshine @DrYohanJohn @NicoleCRust @dsmith @ekmiller @PessoaBrain @kendmiller @bwyble @achristensen56
But in exactly what style does the cerebellum make its predictions? If the cortex learns general associations, the hippocampus learns specific incidents and sequences, and the limbic system specialises in rewards and goals, what extra does the cerebellum do? and in what area?
Something to do with timing??
Cerebellum may only support WM in a distantly foundational way, just as imaging suggests sound-symbol is the basis for abstract semantic processing in a proficient adult reader. In each case, the former might simply be the (still-innervating) neural foundation of the latter.
Myself, I think cerebellum contributes more directly to WM operations, but few are interested in speculations!
@strangetruther @macshine @DrYohanJohn @NicoleCRust @ekmiller @PessoaBrain @kendmiller @bwyble @achristensen56
@dsmith @macshine @DrYohanJohn @NicoleCRust @ekmiller @PessoaBrain @kendmiller @bwyble @achristensen56
I doubt the cerebellum provides symbolism. It's too different structurally to be doing something other parts do with different architecture.
Never be ashamed of speculations! EVERYTHING starts that way!!!
@strangetruther @DrYohanJohn @NicoleCRust @dsmith @ekmiller @PessoaBrain @kendmiller @bwyble @achristensen56 context-dependent timing through a highly multi-dimensional interconnected circuitry. The neuroanatomy is complex, but imho clearly shows how these kinds of capacities emerge. Detailed exposition here if you're interested: https://shine-lab.org/wp-content/uploads/2021/09/2020_progneuro.pdf
@macshine @DrYohanJohn @NicoleCRust @dsmith @ekmiller @PessoaBrain @kendmiller @bwyble @achristensen56
Thanks!! Looks excellent! Probably one of the first I should muse over in detail...
Seems thalamus as grand c-ordinator and normaliser, and cerebellum as doing timing, are supported.
@strangetruther @DrYohanJohn @NicoleCRust @dsmith @ekmiller @PessoaBrain @kendmiller @bwyble @achristensen56 would love to hear any thoughts/notes. In case you're interested, it's kind of the first in a series I've been slowly working on. This is the 2nd one, which incorporates the superior colliculus: https://shine-lab.org/wp-content/uploads/2022/11/2022_nbr.pdf.
Another was just accepted on the role that the neuromodulatory system plays in shaping the balance between the diff subsystems, but no pdf yet. Can share later.
@macshine @DrYohanJohn @NicoleCRust @dsmith @ekmiller @PessoaBrain @kendmiller @bwyble @achristensen56
The abstract of the 1st seems straightforward - promises plenty of useful detail in the paper.
It mentions feed-back / feed-forward; some now say top-down & bottom-up should be preferred?
The meaning of "the distinct thalamic populations" - whether in the Th. or projections from it is no doubt made clear lower down.
So management is another thing Th. took over from Supp. Coll!
@dsmith @DrYohanJohn @ekmiller @PessoaBrain @NicoleCRust @kendmiller @bwyble @achristensen56
This also looks awfully good. Had to use "magic" to get this, and will have to have another go at getting theMiller lab ones.
Already got a week's worth of solid reading from just a couple of days!
@kendmiller @ekmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
Awesome. Such patchiness would be exactly the kind of evidence that I would find convincing. I wonder if it is present in PFC et al.
@bwyble @ekmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
My vague memory is that it was seen in frontal areas. Not a trustworthy memory though. One of us should dig up those old papers. @DrYohanJohn does anatomy, maybe he could identify the papers, or ask Helen Barbas if she could?
I shared our recent perspective paper somewhere else in this thread (I think).
https://www.frontiersin.org/articles/10.3389/fnsys.2022.921468/full
Helen is somewhat skeptical of the column as a canonical structure: M1 is strikingly different from V1, and parts of ACC (such as A25) have almost no granular layer 4. But 'vertical' and 'horizontal' are good organizing principles for local and long-range connectivity.
@kendmiller @bwyble @ekmiller @strangetruther @achristensen56 @NicoleCRust
@DrYohanJohn @bwyble @ekmiller @strangetruther @achristensen56 @NicoleCRust
But do you know about the structure of within-area long-range horizontal connections in motor and frontal cortex? My memory is that these projections are periodic/patchy, but I am not remembering precisely what the paper(s) were or what areas they studied.
@DrYohanJohn @bwyble @ekmiller @strangetruther @achristensen56 @NicoleCRust
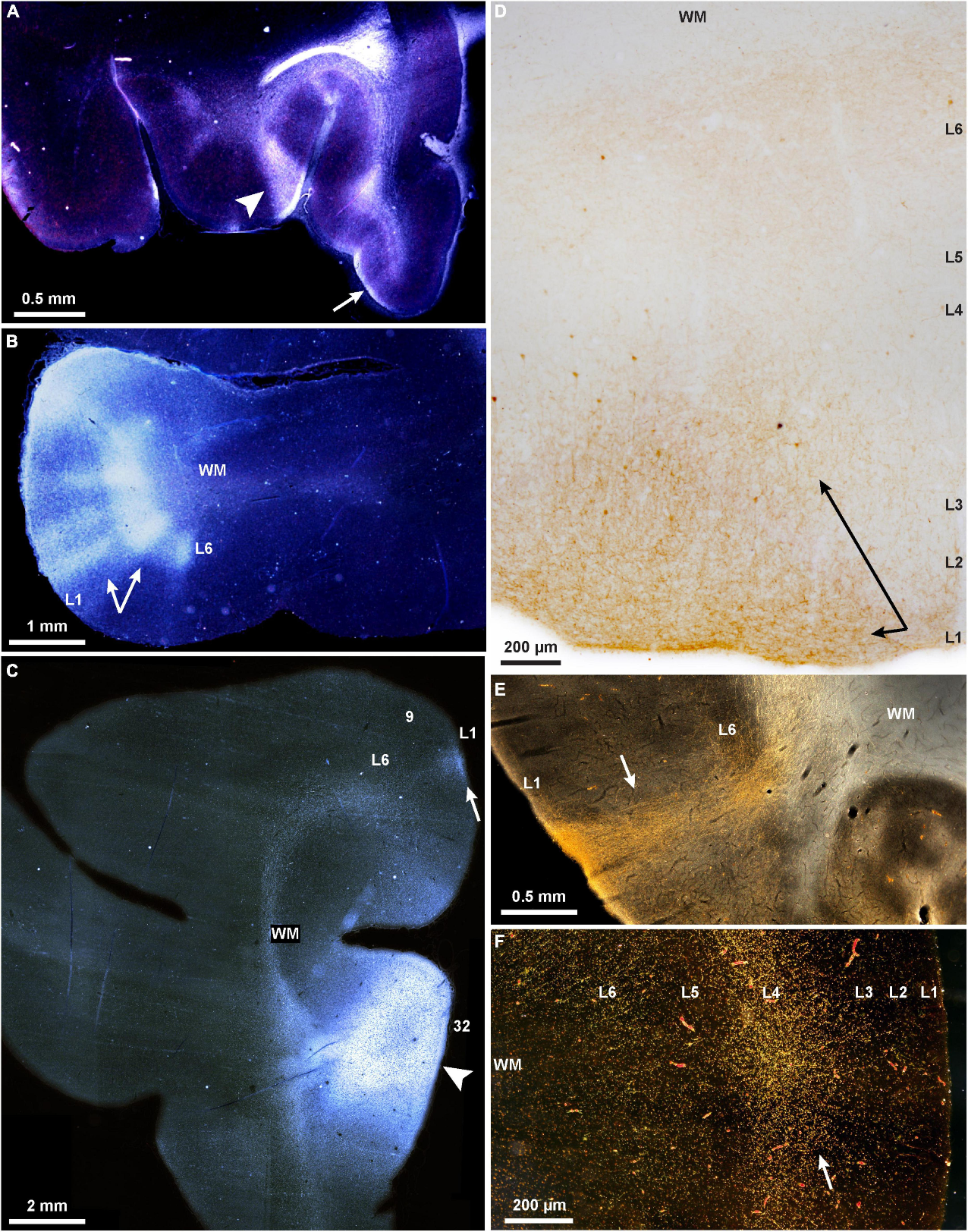
OK here's one example, from Jenny Lund and others, https://onlinelibrary.wiley.com/doi/abs/10.1002/(SICI)1096-9861(19961223)376:4%3C614::AID-CNE9%3E3.0.CO;2-4
The did both anterograde and retrograde tracing from injections in prefrontal areas 9 and 46. Both intrinsic (within-area) projections and "associational" (to other pre-frontal areas) projections formed periodic patches or stripes, with a repeat spacing of about 780 microns for intrinsic connections; no significant differences were seen between structure/distribution of intrinsic vs associational projections, but they seem to have less data on associational and don't specifically state a repeat distance for them.
I attach an example figure. The figure legend reads "Fig. 6. Brightfield photomicrograph of four discrete intrinsic clusters of CtB-positive neurons (arrows) [KM: CTB is a retrograde tracer; they also did the same experiments with the same results with an anterograde tracer] distributed along the dorsal bank of the principal sulcus (PSI in area 46. All clusters arise from a single injection site near the area 9/46 border, which is located immediately to the left (medial) of the left edge of the micrograph. Clusters of anterogradely-labeled axons are also apparent coincident with neuron clusters. Scalebar = 200microns".
Check out Figure 1B. There is a 'columnar' pattern. But the point is also that there are other patterns.
@bwyble @ekmiller @strangetruther @achristensen56 @NicoleCRust
@DrYohanJohn @bwyble @ekmiller @strangetruther @achristensen56 @NicoleCRust
How do you describe that pattern? Periodic but with a weaker continuous pattern across layer 6??
@bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
For example, did you know that there is information in the *electric fields* created by the brain? Further, the electric fields show little or no representational drift, i.e., they are more robust souces of info than the (variable) underlying activity.
https://doi.org/10.1016/j.neuroimage.2022.119058
Brain function is like an orchestra. The individual players (neurons) come and go, but the song (an emergent property) plays on.
@bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
Coming soon: New papers showing that it also works the other way around. LFPs and electric fields driving spiking through ephaptic coupling.
Whoa. If true, that would be a Very Big Deal. Maybe Kuhnian?
@ekmiller @bwyble @kendmiller @strangetruther @achristensen56 @DrYohanJohn
Biophysically speaking it must be possible. I guess the empirical question is whether ephaptic coupling can strongly alter firing when the synaptic input is large.
@NicoleCRust @ekmiller @bwyble @kendmiller @strangetruther @achristensen56

@DrYohanJohn @NicoleCRust @ekmiller @bwyble @kendmiller @strangetruther @achristensen56
How can it not? If physically there aren’t enough ions for all neurons to fire simultaneously, then ephaptic coupling must be a built-in mechanism that ensures sparsity of firing.
@NicoleCRust @bwyble @kendmiller @strangetruther @achristensen56 @DrYohanJohn
Well, there are other examples of ephaptic coupling in the brain. We wouldn't be the first to show it.
But yes, Kuhn knew what he was talking about. People do buy into and defend paradigms. No one know enough about the brain to call anything an epiphenomenon. If we can measure it with our crude instruments, it possible and probable that the brain can use it.
@ekmiller @kendmiller @strangetruther @achristensen56 @NicoleCRust
@DrYohanJohn
Yep, neurons live in a bath of electric fields. Way back in my grad school days we were studying the effect of theta phase on evoked potentials and it was very challenging to separate the contribution from putative cholinergic modulation vs just a change in electrical field. IIRC we concluded that the the effect was at least partially neuromodulatory because the size of the modulation was larger than could be explained by the extracellular voltage swings.
I think this idea is really interesting, and would be a way to modulate activity in brain areas, acting to either dampen or excite. Do you think it also has the ability to communicate substantial amounts of information?
@bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
One idea is that they have the ability to route information around the brain and enable top-down control.
https://doi.org/10.1073/pnas.2014868117
https://doi.org/10.1016/j.neuron.2018.09.023
And there is the work by Pascal Fries and colleagues.
Even sustained attention isn't sustained. It cycles at 4 Hz in sync with LFP/EEG.
https://www.sciencedirect.com/science/article/pii/S0896627318306305
@ekmiller @bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn
How interesting. Like the higher-order manifestation of the changing subset of recurrently connected neurons that can represent information:
Druckmann S, Chklovskii DB. Neuronal circuits underlying persistent representations despite time varying activity. Current Biology. 2012 Nov 20;22(22):2095-103. https://www.sciencedirect.com/science/article/pii/S0960982212010810
#neuroscience
I love this paper. And the amazing experimental work from the Svoboda lab around dynamical systems and short term memory that followed from it. I convinced the very humble Shaul Druckmann to summarize it in a recent paper we wrote:
This work was predated by the understanding that a particular brain area known as Anterior Lateral Motor cortex (ALM) is a key node in short term memory representations of action preparation in mice (Guo et al., 2014). The most central question was: how do the neural dynamics in this circuit support short term memory? We found evidence that persistent activity in ALM supports short term memory, but at the same time these dynamics depend not just on circuits confined to ALM, but are supported instead by a multi-regional network of interactions that operate as complex, dynamical system (Li et al., 2016). Foundational to this discovery was our development of a theoretical framework in which the population dynamics of a brain area are not viewed as a single entity (as is typically the case), but instead as a set of overlaid dynamical patterns, or factors, each with their own computational and functional meaning (Druckmann & Chklovskii, 2010, 2012). This framework allowed us to interpret the results of otherwise perplexing optogenetic brain perturbation experiments. There we determined that persistent memory activity remains robust to perturbations along the dimensions, or patterns, that are linked to short term memory, but not along other dimensions (despite the fact that these other dimensions capture substantial variance in the population response). We also determined that these short term memory representations (and behavior) were surprisingly robust to perturbation as a consequence of distributed interactions between ALM across the two hemispheres (Li et al., 2016). In sum, our understanding of how persistent memory activity arises in ALM depended crucially on the tools we had developed to think about population dynamics along multiple dimensions, and our development of those tools predated a detailed understanding of what we would ultimately use them for.
https://pubmed.ncbi.nlm.nih.gov/36473096/
@albertcardona @ekmiller @bwyble @kendmiller @strangetruther @achristensen56 @DrYohanJohn
@NicoleCRust @ekmiller @bwyble @kendmiller @strangetruther @achristensen56 @DrYohanJohn
At Janelia I saw Shaul and Mitya in action, describing their theoretical findings with amusing toy videos to illustrate how an invariant representation could be held by a variedly active population of recurrently connected neurons. Like projecting a 3D vector onto a 2D surface, there are many combinations of 3D vectors whose combination has the same 2D projection on that surface. Intuitive explanation!
Yup! I like to think of such representations as a relay race. Or 'passing the parcel'.
@albertcardona @NicoleCRust @ekmiller @bwyble @kendmiller @strangetruther @achristensen56

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
@ekmiller @bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn @cogneurophys Ooh - interesting paper alert! of course
1. variable is not the same thing as indecipherable or random, so, the case where LFP is stable may be a more easily-interpretable exception to a more diverse set of drift 'rules',
2. lumpers and splitters can both be happy
--from still-relevant anatomical work of Guy Elston showing early sensory nctx organization gets incrementally "richer" with increasing dist. from sensory thal, (aka 'association' ctx incl dlPFC) and is proportionately amplified across primate species' with increasing brain size
--to Hilgetag Goulas Barbas (and @DrYohanJohn !) changing motifs where morphological/cytoarchitectonics parallel macroscale connectomics. While these anatomical studies do not argue for a single canonical (straw man) column, they suggest specific dimensions of gradients that may influence how a chunk of tissue computes. And computes differently from a chunk at another spot along those dimensions. #neuroethology #neuroanatomy #sizematters
Yup! What you say about motifs and architectonics is precisely what I was trying to say earlier: adding systematic gradients is like adding just one additional term to the 'series approximation'.
@ekmiller @bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @cogneurophys
@DrYohanJohn @ekmiller @bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @cogneurophys Yes! with the caveat that there may be some more discrete 'jumps' than the term gradient implies. e.g. differences in cell type. Take, for example, the 'LR' excit. cell that forms the namesake of granular (not granule cell!) RSC. It creates the conditions for a cortical area w/ qualitative change to its dRSC neighbor. https://www.cell.com/cell-reports/fulltext/S2211-1247(19)31758-9?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS2211124719317589%3Fshowall%3Dtrue
@karihoffman @bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn @cogneurophys
Great point. That is the true true. Just because we don't know the source doesn't mean it is noise.
However, there is like a gazzillion neurons in the brain. Organization of processing can't happen at the neuron-by-neuron level. It must be at a larger scale. There is where LFPs and electric fields come in.
@karihoffman @bwyble @kendmiller @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn @cogneurophys
Here is a preprint from Mikael Lundqvist that makes this point.
Spatial computing for the control of working memory
https://www.biorxiv.org/content/10.1101/2020.12.30.424833v2
Sensory information is fine-grained and repeated across networks. Top-down control is at a maco level as alpha/beta rhythms organize activity on the scale of millions of neuron.
The conductor conducts orchestra sections, not individual players.

@kendmiller @bwyble @strangetruther @achristensen56 @NicoleCRust @DrYohanJohn wouldn't it be too redundant if so many neurons in V1 are doing the same simple job of telling a few not-so-sharp directions... Maybe V1 does many other important things we don't know much yet...